Quelques-uns des principaux sujets de Géoforum

▲ Vente et achat de minéraux français et cristaux du monde sur Internet ▲ |

Vente et achat de minéraux sur Internet
Voir le nouveau contenu de Géoforum


gaeldeploeg
-
Compteur de contenus
6895 -
Inscription
Type de contenu
Profils
Forums
Galerie
Blogs
Boutique
Tout ce qui a été posté par gaeldeploeg
-
 Bonjours BelgoRaptor! Quel genre de paléontologie aime tu, car il y en a des centaines de façon de faire de la paléonto. Perso, je la pratique en amateur pour ne pas avoir a faire de rapport d'activité ou remplir des tonnes de formulaires administratifs. en +, çà permet de ne pas altéré la passion
Bonjours BelgoRaptor! Quel genre de paléontologie aime tu, car il y en a des centaines de façon de faire de la paléonto. Perso, je la pratique en amateur pour ne pas avoir a faire de rapport d'activité ou remplir des tonnes de formulaires administratifs. en +, çà permet de ne pas altéré la passion -
Les dinosaures bannis des écoles de New York (créationisme)
gaeldeploeg a répondu à un sujet de Théophraste dans Forum Fossiles et Paléontologie
http://kinea.media/fr/science-technologie/quatre-erreurs-sur-l-evolution L'intégrale du lien. pourquoi? par ce que les liens s'altères avec le temps! Quatre erreurs sur l'évolution La controverse qui a débuté en 1859 concernant le concept de la sélection naturelle décrit dans le livre de Charles Darwin De l'Origine des Espèces n'a jamais vraiment cessé. Plus que jamais, des mouvements religieux militent aux États-Unis, en Turquie et dans d'autres pays, pour que soit enseignée en cours de Biologie, la thèse du dessein intelligent (intelligent design) comme explication alternative du fonctionnement de la vie. Les études d’opinion confirment bien d’ailleurs que les thèses créationnistes ou apparentées ont un large écho aux États-Unis1. Cette défiance est expliquée comme résultant de plusieurs facteurs, dont la mauvaise compréhension qu’a le public de la théorie de l’évolution et de ses mécanismes. Dans les débats, même les tenants d’un enseignement exclusif de l’évolution en cours de Biologie semblent bien souvent peu familiers avec ce qu’ils défendent2. Pourtant, l’évolution est l’un des concepts scientifiques les plus abondamment vulgarisés ; il a paradoxalement subi les effets pervers de cette popularisation : au fil des décennies s’est développée dans le grand public une suite d’idées sur ce qu’était l’évolution ; et ces idées se sont mues très rapidement en préconceptions profondément inculquées chez le jeune citoyen. Dès le plus jeune âge, nous sommes influencés par les références culturelles telles que les films, publicités, livres, journaux, débats qui nous assènent subtilement des contre-vérités concernant cette théorie. Nous allons jeter un coup d’œil à quatre de ces préconceptions largement répandues3. Une explication simplifiée de la sélection naturelle. La version incorrecte correspond à une idée largement répandue de la sélection naturelle : les espèces y sont vues comme uniformes et comme réagissant aux changements environnementaux ; les mutations qu'elles subissent seraient exclusivement dirigées vers l'amélioration. En réalité, il vaudrait mieux considérer la sélection naturelle comme le résultat de deux phénomènes : des mutations non orientées et de la sélection non aléatoire. Les mutations sur le génome des gamètes sont la cause de nouveaux traits non orientés chez les individus qui en résultent (cerclés de bleu sur le schéma) ; les mutations aléatoires sont responsables de la variabilité au sein de la population. La sélection non aléatoire est le processus par lequel est déterminé si ces traits aident ou non le porteur à avoir une plus large progéniture que ses congénères. Les individus qui ne sont pas adaptés à leur environnement n'auront peu ou pas de descendance (ils sont indiqués par des croix rouges), alors que les autres diffuseront leurs traits dans la population. Ici, la population n'est jamais complètement homogène et des mutations apparaissent aléatoirement, mais seules celles qui sont avantageuses sont sélectionnées. Ce schéma est inspiré de (Gregory, 2009). Il n'y a pas de hiérarchie du vivant Habituellement, on décrit l'évolution comme une sorte de hiérarchie du vivant, de laquelle les organismes les plus simples (les bactéries) seraient à la base et les plus complexes (l’être humain) seraient au sommet. En ce sens, l'évolution serait allée de la bactérie, à l'animal unicellulaire, au poisson, au mammifère pour finalement déboucher sur l'être humain ; du niveau le plus bas au plus haut, du plus simple au plus complexe. Mais cette vision des choses ne tient pas quand on essaye de remplir de manière exhaustive cette hiérarchie du vivant : par exemple, comment déterminer l'animal le plus "élevé" sur cette échelle entre le lion et l'éléphant, ou entre les fourmis et les abeilles ? En fait, l'évolution ne décrit pas une hiérarchie du vivant, mais plutôt comment les animaux actuels sont apparentés. La sélection naturelle, le principal mécanisme de l'évolution4, stipule que tous les individus qui existent aujourd’hui sont issus de lignées qui ont réussi à transmettre leur patrimoine génétique à travers les générations. D'innombrables stratégies pour transmettre l'héritage génétique ont émergé au cours du temps et tous les organismes actuels ont la leur5. En ce sens, il est difficile de qualifier les bactéries comme « inférieures » : elles ont réussi, aussi bien que d'autres organismes, à se reproduire jusqu'à nos jours. Tous les organismes vivants, des bactéries aux humains, en passant par les chênes et les fourmis, sont sur la même « marche » de l’évolution : ceux qui sont toujours là et qui continuent à se reproduire encore aujourd’hui. L’évolution n’est pas « allée du plus simple au plus complexe », ou « allée de l'inférieur au supérieur », elle est allée de l’organisme ancestral à l’ensemble des organismes actuels. Illustration par Renaud Helbig L’idée d’une hiérarchie de la nature où l’humain est au sommet est une idée profondément implantée dans la pensée occidentale bien avant que la notion d'évolution n’apparaisse : Aristote usait déjà d'une telle scala naturæ dans De l'âme, Carl Linnaeus aussi quand il développa sa classification dans Systema Naturæ, elle transparaît encore chez le défenseur et ami de Darwin, Alfred Russel Wallace6. Pour les personnes faisant une telle classification, la force ou l'intelligence de l'animal seraient de bons critères pour ranger les êtres vivants. Mais nous allons voir que ces caractéristiques ne sont pas tout le temps aussi cruciales du point de vue de la sélection naturelle. La "loi du plus fort" n'est pas une loi de la nature La sélection naturelle ne décrit-elle pas une lutte impitoyable entre individus pour pouvoir transmettre ses gènes ? Dans ce contexte n’est-il pas logique de penser que l’individu le plus fort sera capable de tuer sa proie, de menacer le prédateur et de vaincre les rivaux pour la reproduction. Être le plus fort semble être le meilleur moyen de devenir "le gagnant de l'Évolution" avec à la clé, plus de nourriture, moins de danger et aucune concurrence sexuelle, tout ce dont on a besoin pour avoir une large descendance. Le problème est qu'habituellement être fort a un coût, et ce coût n'en vaut pas les avantages dans tous les contextes. Face à un prédateur, il peut être préférable de fuir rapidement ou d'être petit pour pouvoir se cacher facilement, plutôt que d'affronter directement la menace ; en cas de famine, il vaut mieux avoir stocké de la graisse que d'avoir produit une grande masse musculaire. En fait la sélection naturelle ne s'oriente pas dans la même direction quelque soit le contexte : tout individu est l'aboutissement d'une sélection qui le rend adapté à un environnement particulier (qui inclut le climat, les ressources, mais aussi ses congénères et les espèces environnantes). Si au sein de cet environnement, être le plus fort pour pouvoir dominer les autres est un avantage reproductif, alors sans nul doute la sélection naturelle privilégiera ce caractère. Sauf que la sélection naturelle ne se résume pas à un duel physique entre individus ; au contraire, bien souvent, le bénéfice revient plutôt à l'individu discret, parcimonieux ou prudent. Les êtres aussi faibles que sont les souris ou les moineaux ont eux aussi été sélectionnés au cours des temps géologiques ; s’ils sont là aujourd’hui, ce n’est pas à cause de quelque soi-disant "erreur de la nature", mais bien parce que dans leur contexte évolutif, être le plus fort n’est pas un avantage. Donc la loi du plus fort ne s'applique pas à la nature, car il y a d'autres caractéristiques qui, en dépendant de l'environnement, peuvent vous donner de plus grandes chances de survie. Ce qui importe donc alors, c'est d'avoir des caractéristiques qui permettent aux individus de survivre, pas vrai ? Eh bien, en fait... Pas tout à fait... La survie n'est pas le but final de la sélection naturelle Être capable de survivre semble être une caractéristique clé de la nature. Il est frappant de voir que l'aptitude à la survie est universellement partagée par l'ensemble des êtres vivants : un animal peut dépenser toute son énergie à fuir un prédateur, un arbre peut renaître de ses racines, une bactérie peut répondre à une menace antibiotique, etc. D'après cette simple observation, il semble logique d'affirmer que la survie est le but ultime du point de vue de la sélection naturelle. Pourtant, certains cas de mort délibérée peuvent être observés dans la nature : chez certaines espèces d’araignées, les mères se laissent mourir auprès de leurs rejetons pour pouvoir enfin… être mangées par eux !7 Ce comportement est un moyen de donner à la progéniture un bon départ dans la vie. De manière plus connue, une abeille ouvrière n’hésitera pas à se sacrifier en piquant l’animal qui s’approcherait trop de la ruche, et ce simplement pour pouvoir protéger ses sœurs aptes à se reproduire (dans une ruche, toutes les abeilles sont sœurs) - sœurs qui possèdent donc une large part de leur patrimoine génétique. En fait, même si la survie est un instinct qui a été largement sélection au cours de l'évolution, elle n'en est pas pour autant le principe premier de la sélection naturelle, mais juste l'une de ses conséquences. Survivre est certes une condition sine qua non pour se reproduire : un animal qui survit jusqu’au moment propice à sa reproduction a davantage de chances de voir ses gènes transmis aux générations futures. Mais au-delà de la reproduction, si l’individu est plus utile mort que vivant pour la transmission du patrimoine génétique, l’évolution n’a pas d’état d’âme pour le sacrifier sur l’autel de la sélection naturelle. La question n'est pas tant de savoir si l'individu est apte à survivre, mais plutôt si cette survie contribue d'une quelconque manière à avoir de plus grandes chances de transmettre ses gènes. La sélection naturelle ne fait pas agir l'individu pour le bien de son espèce Reprenons l'exemple des abeilles qui se sacrifient. Il s'agit là d'un effet où l'insecte semble avoir un comportement purement altruiste, où son action lui est délétère (dans ce cas précis, il en meurt), et ne bénéficie qu'à ses congénères. La coopération, comme pour des lionnes chassant en groupe, est un autre comportement qui semble indiquer que l'individu a une tendance naturelle à aider ses semblables. À la fois l'altruisme et la coopération semblent tout à fait compréhensible si l'on considère qu'une espèce est un groupe solidaire, qui va lutter contre des menaces communes, souvent représentées par les autres espèces : les abeilles contre les frelons, les lions contre les éléphants, etc. On peut tout d'abord noter que la notion d’espèce est très délicate à manier, même pour des spécialistes. Par exemple, si vous pensez qu’une espèce est un groupement d’individus qui peuvent se reproduire entre eux (comme on nous l’explique habituellement en cours de Biologie), vous laissez de côté la majorité du vivant : les bactéries, levures et autres organismes unicellulaires qui se reproduisent de manière asexuée.8 Mais, encore plus important, il y a le problème des individus non coopératifs. Prenons un exemple : dans une colonie bactérienne, l'essentiel des cellules semblent altruistes. Elles produisent et diffusent dans le milieu une protéine complexe qui permet de consommer une ressource. Si le produit n'est pas directement bénéfique pour la cellule qui l'a produite, mais aide toutes les cellules, et ce (et ceci est important) sans aucune distinction, alors ce comportement peut être considéré comme purement altruiste : les cellules utilisent des ressources pour produire quelque chose que tout le monde peut utiliser. Il semblerait alors dans ce cas que les bactéries ont une tendance naturelle à aider les membres de leur espèce. Mais cette explication simple ne tient pas la route : imaginons que l'une des bactéries a une mutation qui l'empêche de produire cette protéine particulière, elle pourrait alors bénéficier de l'altruisme des autres (en pouvant consommer la ressource) sans pour autant payer le coût de la production de la protéine. Ce gain en énergie peut être investi dans sa reproduction. D'après la sélection naturelle, après quelques générations, cette mutation se sera largement diffusée au sein de la population et le comportement altruiste sera en déclin. Avec cet exemple, on montre que des explications du type "les individus ont tendance à agir pour le bien de leur espèce" ne permettent pas d'expliquer les comportements altruistes ou coopératifs chez les êtres vivants ; nous avons besoin d'explications bien plus subtiles. Pour les précédents exemples, les biologistes de l'évolution expliquent habituellement l'émergence de ces comportements par des arguments où seul l'intérêt purement personnel est considéré : le but est toujours de mieux transmettre ses gènes que ses congénères. Les abeilles qui se sacrifient en piquant le prédateur ne le font pas pour toutes les abeilles du monde, mais uniquement pour leurs sœurs de leur ruche avec qui elles partagent l'essentiel de leur patrimoine génétique9. Si quelques animaux chassent ensemble, ce n'est pas par solidarité pour l'espèce, mais simplement parce que plus ils sont nombreux, plus grosses sont les proies potentielles. Si la part de chaque chasseur est plus grande que ce qu'il aurait eu en chassant seul, alors il est dans son intérêt égoïste de chasser en groupe.10 Conclusion À mon sens, si les principes de la sélection naturelle ont tellement de mal à passer dans le grand public, c’est qu’ils vont à l’encontre de certaines idées profondes que l’on se fait sur la vie et sur l’être humain. On aimerait se voir comme au sommet du règne de la vie, comme étant nécessairement le résultat d’une sélection poussant à être toujours plus fort et toujours plus intelligent, ou comme faisant naturellement partie d’un groupe solidaire qui fait face ensemble à l’adversité. Malheureusement, ou heureusement, la science ne suit pas toujours nos aspirations philosophiques. Elle n'est pas là pour nous plaire ou pour flatter notre ego, mais simplement pour nous donner une vue cohérente de ce qu'est le monde. Bibliographie Alters, B.J., Nelson, C.E., and Mitton, J. (2002). Perspective: teaching evolution in higher education. Evolution 56, 1891 1901. Clark, C.W., and Mangel, M. (1986). The evolutionary advantages of group foraging. Theoretical Population Biology 30, 45–75. Dawkins, R. (1976). The Selfish Gene (Oxford University Press, USA). Evans, T.A., Wallis, E.J., and Elgar, M.A. (1995). Making a meal of mother. Nature 376, 299 299. Gregory, T.R. (2008). Understanding Evolutionary Trees. Evo Edu Outreach 1, 121 137. Gregory, T.R. (2009). Understanding Natural Selection: Essential Concepts and Common Misconceptions. Evo Edu Outreach 2, 156 175. Gregory, T.R., and Ellis, C.A.J. (2009). Conceptions of Evolution among Science Graduate Students. BioScience 59, 792 799. (Alters et al., 2002) ↩ (Gregory and Ellis, 2009) ↩ Il en existe de nombreuses autres ; d'autres idées reçues peuvent être trouvées sur le site http://evolution.berkeley.edu/evolibrary/misconceptions_faq.php (en anglais) ↩ Il y a d'autres mécanismes existants pour la diffusion de nouveaux caractères tels que la dérive génétique ou la sélection sexuelle, etc. ; je ne traiterai ici que de la sélection naturelle. ↩ Voir l'article de Agustín Ávila sur Kinea: http://kinea.media/fr/science-technologie/agave-chauve-souris ↩ Voir le chapitre XV de Wallace, A.R. (1889). Darwinism, an exposition of the theory of natural selection with some of its applications. ↩ (Evans et al., 1995) ↩ Charles Darwin était au courant des difficultés pour définir clairement la notion d'espèce. Il consacre la totalité du huitième chapitre de son livre à ce propos (Darwin, 1859). ↩ Il s'agit là de la sélection de parentèle. Voir par exemple (Dawkins, 1976) ↩ En réalité, cet exemple est encore plus riche en termes de la sélection naturelle. Voir (Clark and Mangel, 1986). ↩ Renaud Helbig 2016-01-14 Édité par Agustín Ávila Casanueva et Annalí Casanueva Artís Relu par Théo Zimmermann
-
CLIP "LAISSER PASSER" SHILOM MC Oui, et dans touts ce qu'il fait! comme les argiles et les silex. la kaolinite vient forcément de loin, plus d'une centaine KM.
-
Le spinosaure était un piscivore semi aquatique.... Et mangeur de requin!
gaeldeploeg a répondu à un sujet de Chicxulub dans Dinosaures
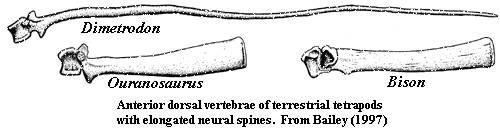
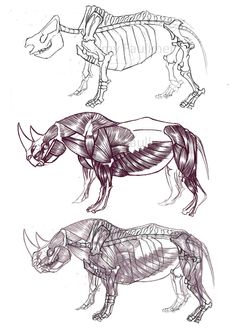
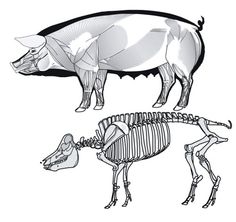
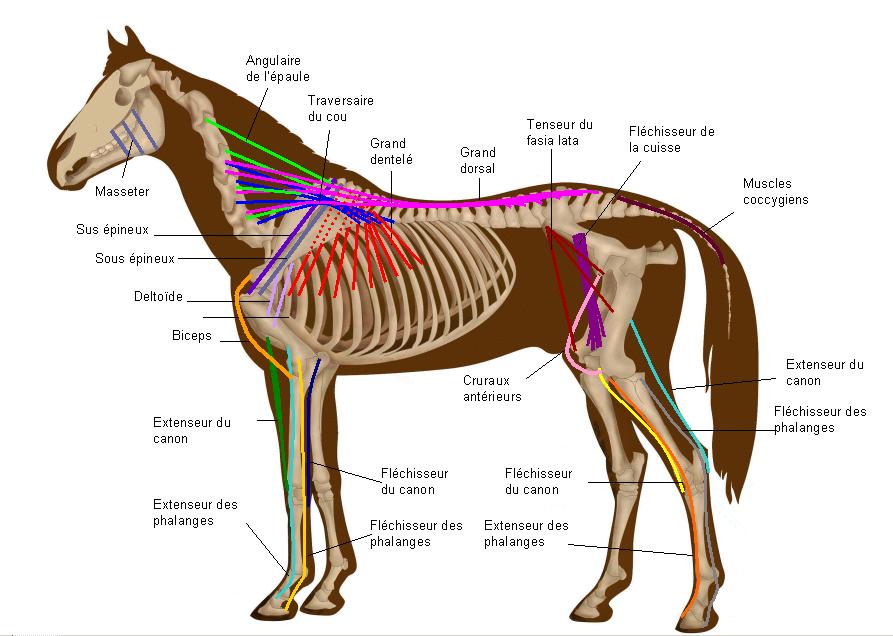
L'Ouranosaure, pour le quel les fossiles sont plus complets. Les épines postérieures été liés ensemble par des tendons ossifiés, une masse musculaire et pas un voile de peau ou une membrane. Un peu de parcimonie, pour soutenir une membrane de peau, des excroissance osseuse moins résistante suffirai. Un peut d'anatomie: Sur les apophise épineuse des vertèbres de dinosaure, observe t'on les points de fixation des ligaments supraspinaux? oui! On cite le bison. Mais il y a d'autres exemples, Les rhinos. Les cochons. Quelle est la fonction des processus épineux chez les vertébrés? d'attache musculaire! Les masses adipeuses non pas besoin d'appuis squelettique! un exemple avec la bosse du chameau. quoi qu'heu. A lire sur le sujet: Tall spines and sailed backs: A survey of sailbacks across time http://reptilis.net/2014/06/21/tall-spines-and-sailed-backs-a-survey-of-sailbacks-across-time/ Angielczyk, K.D., Schmitz, L. 2014. Nocturnality in Synapsids Predates the Origin of Mammals by Over 100 Million Years. Proc. R. Soc. B. Early Access. Angilletta Jr., M.J., Werner, Y.L. 1998. Australian Geckos do not Display Diel Variation in Thermoregulatory Behavior. Copeia. Vol. 3:736–742. Autumn, K., De Nardo, D.F. 1995. Behavioral Thermoregualtion Increases Growth Rate in a Nocturnal Lizard. J. Herp. Vol. 29(2):157–162. Baumel, J.J. 1975. Aves heart and blood vessels. in: Getty, R. (ed). The Anatomy of the Domestic Animals, 5th ed., Vol. 2. Philadelphia. pps: 1968–2009. Dzialowski, E.M., O’Connor, M.P. 1998. Utility of Blood Flow to the Appendages in Physiological Control of Heat Exchange in Reptiles. J. Therm. Biol. Vol. 24:21–32. Dzialowski, E.M., O’Connor, M.P. 2004. Importance of the Limbs in the Physiological Control of Heat Exchange in Iguana iguana and Sceloporus undulatus. J. Therm. Biol. Vol. 29:299–305. Falk, D. 1990. Brain Evolution in Homo: The “Radiator” Theory. Behav. Brain. Sci. Vol. 13:333–381. Heath, J.E. 1964. Head-Body Temperature Differences in Horned Lizards. Phys. Zool. Vol. 37(3):273–279. Heath, J.E. 1966. Venous Shunts in the Cephalic Sinuses of Horned Lizards. Phys. Zool. Vol.39(1):30–35. Rowe, M.F. 2012. Sensory Biophysical Variations in Resting and Exercising Elephants: Energetic, Thermoregulatory, and Behavioral Adaptations. AZ State U. Dissertation. Rommel, S.A., Pabst, D.A., McLellan, W.A. 1998. Reproductive Thermoregulation in Marine Mammals. Am. Sci. Vol. 86:440–448. Seidel, M.R. 1979. The Osteoderms of the American Alligator and their Functional Significance. Herpetologica. Vol. 35(4):375–380. Shoshani, J., Kupsky, W.J., Marchant, G.H. 2006. Elephant Brain Part 1: Gross Morphology, Functions, Comparative Anatomy, and Evolution. Brain. Res. Bull. Vol. 70:124–157. Zippel, K.C., Lillywhite, H.B., Mladinich, R.J. 2003. Anatomy of the Crocodilian Spinal Vein. J. Morph. Vol. 258:327–335.
-
Le spinosaure était un piscivore semi aquatique.... Et mangeur de requin!
gaeldeploeg a répondu à un sujet de Chicxulub dans Dinosaures
C'est le coup de crayon de Mazan http://petitcarnetpaleo.blogspot.fr/2016_10_01_archive.html A bon!?et pourquoi? -
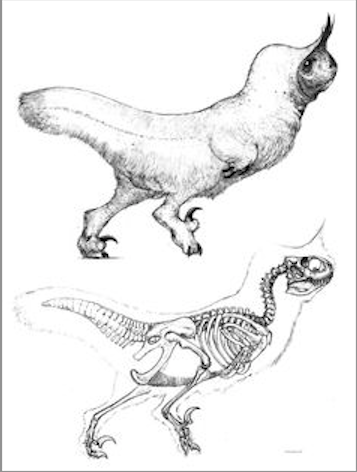
Préservation des tissus cérébraux chez un dinosaure
gaeldeploeg a répondu à un sujet de dorian. dans Dinosaures
Une version amélioré d'un fossile 'classique', le moulage endocrinien. Chaque crâne fossile en contient un normalement, mais il faut cassé le crâne En 2015, pour l'expo temporaire' à vos cerveaux'au Muséum de Brussel.
-
Tentacules. Ben Non, because De plus avec la rigidité cadavérique le corps devait se rétracter dans la loge d'habitation et les gaz dans les loges on pu faire flotté les coquilles long temps. pas d'enfouissement rapide pour les cadavres d'ammonites. voila pour la taphonomie des bèbêtes
-
technique de preparation d'un fossile
gaeldeploeg a répondu à un sujet de ammonite dans Forum Fossiles et Paléontologie
Tools and Preparation Isotelus Preparation STEP by STEP May 30, 2015 | Gianpaolo Dr. Di Silvestro Ricevo mail che mi chiedono se i trilobiti sono veri, come vengono preparati, quali sono i migliori strumenti ecc. Ad uso didattico allego delle sequenze fotografiche che spiegano step by step i processi "di preparazione"; non sono gli unici metodi, ma é il sistema che preferisco. Several customers are sending me mails about preparations, fake trilobite quastions and so on. Well i am posting a Trilobite preparation sequences to show at all one method to prep such specimen. It is not the only one preparation method, it is just one. 1) Individuazione della sezione / checking the fossil section 2) Incollaggio sezioni adiacenti; a tal proposito é necessario usare un buon-ottimo mastice per pietra (evitare marche economiche e scadenti). Uso Tenax solido di varie colorazioni in base alla tipologia di pietra. Non usare sotto temperature inferiori ai 10°C e suepriori ai 30°C nella catalizzazione assicurarsi di rimuovere le sbavature con una taglierina ben affilata. Gluing parts with rock bicomponent glue. I use tenax, it is the best stone glue on Italian market. Usually there are several coloured glue. You can use depending by matrix colours. Don't use it under 10°C or 30°C up. During the catalysis process remove the exceding parts of glue with a blade. 3) Rimozione pneumatica di parte della matrice. Chicago Pneumatic CP9361 tools a 6 Bar / Removing matrix with Chicago pneumatic tools CP9361at 6 Bar power. 4) fasi di incollaggio successive / Other parts fixed 5) Uso di resine epossidiche bicomponenti per la ricostruzione di eventuali parti mancanti. puoi trovarle nel nostro shop online/ Apoxie resin to recontruct missing parts, available on our shop 6) Sabbiatura finale a 25 micron con Ossido di Alluminio / Sand blasting process with 25 micron sand of Alluminium Oxide. Isotelus gigas forma Giovanile Isotelus gigas Ordoviciano Rust Formation, Trenton Group Walcott-Rust Quarry, Trenton Falls, New York Restauro 3% occhio dx 6 hr d preparazione presente nel nostro shop Isotelus gigas in juvenile form Ordoviciano Rust Formation, Trenton Group Walcott-Rust Quarry, Trenton Falls, New York restoration on right eyes at 3% 6 hr preparation available in our shop -
Salut! Là, un trèèèès beau travail! triloblog category Paleoart! çà déchire la 3D. Ou sont les limites? Déjà qu'avec l'impression l'humanité avait fait un bond
-
Nouveau concept : PaleoArt' !
gaeldeploeg a répondu à un sujet de Delaroue dans Forum Fossiles et Paléontologie
triloblog category Paleoart! -
Fossiles et mineraux pres de Nimes
gaeldeploeg a répondu à un sujet de Annes dans Forum Fossiles et Paléontologie
!? Plutôt Durfort-et-Saint-Martin-de-Sossenac. avec son mammouth. Passe par Monfrin. Et la=>atelier la trouvaille -
Echange fossiles contre coquilles d'escargots !
gaeldeploeg a répondu à un sujet de Bathollovien dans Echanges de fossiles
Bonjours! Je connais un talus très riche en restes de gastéropodes terrestres actuels. J'échantillone dans les deux jours. Pour l'échange, justeun jour ou l'autre!? -
La terminologie de la ref don j'ai sortie les quelques feuilles si-dessus est 'THE référence'! "Groupe des résines fossiles: succinctes et rétinites"./ et pas ambre ou copal. « Le succin de la Baltique » et pas ambre . ‘la résine de copal’ et pas le copal et encore moins, un copal, puisque c'est la résine sub-fossile de cet arbre uniquement. ETC, ETC… touts le reste n'est que des"conneries", voir même, C'est de la merde! A touts, Amicalement! G.d.P, ( Granulats de Picardie et/ou Gaël de Ploëg ).
-
Bon survole alors! Mesozoic Climate Constantine, A., A.Chinsamy, T.H.Rich, & P.Vickers-Rich. 1998. Periglacial environments and polar dinosaurs. South African Journal of Science 94:137-141. Douglas, J.G., & G.E.Williams. 1982. Southern polar forests: The Early Cretaceous floras of Victoria and their palaeoclimatic significance. Palaeogeography, Palaeoclimatology, Palaeoecology39:171-185. Frakes, L.A & J.E.Francis 1988 A guide to Phanerozoic cold polar climates from high-latitude ice-rafting in the Cretaceous. Nature 333:547-549 Frakes, L.A., Alley & Deynoux 1995 Early Cretaceous Ice Rafting and Climate Zonation in Australia. International Geology Review 37:567-583 Gregory, R.T., Douthitt, C.B., Duddy, I.R., Rich, P.V. and Rich, T.H. 1989. Oxygen isotopic composition of carbonate concretions from the Lower Cretaceous of Victoria, Australia. Implications for the evolution of meteoroic waters on the Australian continent in a paleopolar environment. Earth and Planetary Science Letters 92: 27-42. Parrish, J.T., R.A.Spicer, J.G.Douglas, T.H.Rich, & P.Vickers-Rich. 1991. Continental climate near the Albian South Pole and comparison with climate near the North Pole. Geological Society of America, Abstracts with Programs 23:A302. Rich, P.V., Rich, T.H., Wagstaff, B.E., McEwen-Mason, J., Douthitt, C.B., Gregory, R.T. and Felton, E.A. 1988. Evidence for low temperatures and biologic diversity in Cretaceous high latitudes of Australia. Science 242:1403-1406. Rich, T.H., Rich, P.V., Wagstaff, B., McEwen-Mason, J., Douthitt, C.B. and Gregory, R.T. 1989. Early Cretaceous biota from the northern side of the Australo-Antarctic Rift Valley. In J.A.Crame (ed.) Origins and Evolution of the Antarctic Biota. Geological Society Special Publication 47:121-130. Wagstaff, B.E., & J.McEwen-Mason. 1989. Palynological dating of lower Cretaceous coastal vertebrate localities, Victoria, Australia. National Geographic Research 5:54-63. References Dinosaurs, plesiosaurs, mosasaurs and more: New light on New Zealand Mesozoic reptiles By R Ewan Fordyce The text below is modified slightly from an original review which appeared as: Fordyce, R. E. 2006. New light on New Zealand Mesozoic reptiles. Geological Society of New Zealand newsletter 140: 6-15. Thanks to the President and Editor of the Geological Society for permission to post this review. You can get a pdf of the article. ou http://www.otago.ac.nz/geology/research/paleontology/mesozoic-reptiles.html Brochu, C. A. 2001. Progress and future directions in archosaur phylogenetics. Journal of Paleontology 75 (6): 1185-1201. Caldwell, M. W., Holmes, R., Bell, G. L., & Wiffen, J. 2005. An unusual tylosaurine mosasaur from New Zealand: A new skull of Taniwhasaurus oweni (lower Haumurian; upper Cretaceous). Journal of Vertebrate Paleontology 25 (2): 393-401. Callaway, J. M. & Nicholls, E. L. (eds) 1997. Ancient Marine Reptiles. Academic Press, San Diego. 448 p. Campbell, H. J., Andrews, P. B., Beu, A. G., Maxwell, P. A., Edwards, A. R., Laird, M. G., Hornibrook, N. de B., Mildenhall, D. C., Watters, W. A., Buckeridge, J. S., Lee, D. E., Strong, C. P., Wilson, G. J., & Hayward, B. W. 1993. Cretaceous-Cenozoic geology and biostratigraphy of the Chatham Islands, New Zealand. Institute of Geological and Nuclear Sciences monograph 2: 1-269. Campbell, J. D. 1965. New Zealand Triassic saurians. NZ journal of geology and geophysics 8 (3): 505-509. Cooper, R. A. (ed.) 2004. The New Zealand geological timescale. Institute of Geological and Nuclear Sciences monograph 22: 1-284. Cruickshank, A. R. I. & Fordyce, R. E. 2002. A new marine reptile (Sauropterygia) from New Zealand: further evidence for a Late Cretaceous austral radiation of cryptoclidid plesiosaurs. Palaeontology 45 (3): 557-575. Farlow, J. O. & Brett-Surman, M. K. 1997. (eds), The complete dinosaur. Indiana University Press, Bloomington. 752 p. Fleming, C. A. 1962. New Zealand biogeography. A paleontologist's approach. Tuatara 10: 53-108. Fleming, C. A., Gregg, D. R., & Welles, S. P. 1971. New Zealand ichthyosaurs - a summary, including new records from the Cretaceous. NZ journal of geology and geophysics 14: 734-741. Fordyce, R. E. 2003. Fossils and the history of life. Pp 35-64 in Darby, J. T., Fordyce, R. E., Mark, A. F., Probert, P. K., & Townsend, C. R. (eds), The natural history of southern New Zealand. University of Otago Press, Dunedin. 387 p. Fordyce, R. E., Campbell, H. J., and Warren, A. 2003. A presumed stereospondyl (Amphibia, Stereospondyli) from the marine Triassic of Titiroa Stream, Mataura Valley, Southland, New Zealand. Journal of the Royal Society of NZ 33 (1): 301-306. Gasparini, Z., et al. 2003. The elasmosaurid plesiosaur Aristonectes Cabrera from the latest Cretaceous of South America and Antarctica. Journal of Vertebrate Paleontology 23 (1): 104-115. Gregg, D. R. 1997. Sam Welles in New Zealand. Geological Society of New Zealand newsletter 114: 95-97. Haast, J., 1870. Notes on a collection of saurian remains from the Waipara River, Canterbury, in the possession of J.H. Cockburn Hood, Esq. Transactions and proceedings of the NZ Institute 2: 186-189. Haw, D. 2002. The discovery of fossils in Mangahouanga Stream. Geological Society of New Zealand newsletter 129: 13-14. Hector, J. 1874. On the fossil Reptilia of New Zealand. Transactions of the NZ Institute 6: 333-358. Hiller, N. & Mannering, A. A. 2004. Elasmosaur (Reptilia: Plesiosauria) skull remains from the Upper Cretaceous of North Canterbury, New Zealand. Records of the Canterbury Museum 18: 1-7. Hiller, N. & Mannering, A. A. 2005. An unusual new elasmosaurid plesiosaur (Sauropterygia) from the Upper Haumurian (Maastrichtian) of the South Island, New Zealand. Memoirs of the Queensland Museum 51 (1): 27-37. Hiller, N., Mannering, A. A., Jones, C. M., & Cruickshank, A. R. I. 2005. The nature of Mauisaurus haasti Hector, 1874 (Reptilia: Plesiosauria). Journal of Vertebrate Paleontology 25 (3): 588-601. King, P. R. 1998. Paleogeographic reconstructions of New Zealand. Geological Society of NZ miscellaneous publication [Geology and genes symposium] 97: 45-48. Lawver, L. A., Gahagan, L. M., & Coffin, M. F. 1992. The development of paleoseaways around Antarctica. American Geophysical Union, Antarctic research series 56: 7-30. Long, J. A. 1998. Dinosaurs of Australia and New Zealand, and other animals of the Mesozoic Era. University of New South Wales, Sydney. 188 p. McKay, A., 1877. Oamaru and Waitaki districts. NZ Geological Survey report of geological explorations 1876-77, (10): 41-66. McKee, J. W. A. & Wiffen, J. 1998. Mangahouanga Stream: New Zealand's Cretaceous dinosaur and marine reptile site [field guide]. Geological Society of NZ miscellaneous publication 96: 1-18. McLoughlin, S. 2001. The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism. Australian Journal of Botany 49 (3): 271-300. Meek, E. P. 1991. Sedimentology of two formations within the Tioriori Group, Chatham Island. BSc Hons thesis, Department of Geological Sciences, University of Canterbury, Christchurch. 86 p. Molnar, R. E. 1981. A dinosaur from New Zealand. Pages 91-96 in Cresswell, M. and Vella P. (eds), Fifth International Gondwana Symposium, Wellington, New Zealand, 11-16 February 1980. Balkema, Rotterdam. Molnar, R. E. & Wiffen, J. 1994. A Late Cretaceous polar dinosaur fauna from New Zealand. Cretaceous Research 15 (6): 689-706. Molnar, R. E., Wiffen, J., and Hayes, B. 1998. A probable theropod bone from the latest Jurassic of New Zealand. NZ Journal of Geology and Geophysics 41 (2): 145-148. O'Keefe, E. R. 2004. On the cranial anatomy of the polycotylid plesiosaurs, including new material of Polycotylus latipinnis, Cope, from Alabama. Journal of Vertebrate Paleontology 24 (2): 326-340. Pole, M. S. 1999. Structure of a near-polar latitude forest from the New Zealand Jurassic. Palaeogeography Palaeoclimatology Palaeoecology 147 (1-2): 121-139. Reguero, M. A., Marenssi, S. A., & Santillana, S. N. 2002. Antarctic Peninsula and South America (Patagonia) Paleogene terrestrial faunas and environments: biogeographic relationships. Palaeogeography Palaeoclimatology Palaeoecology (3-4): 189-210. Rich, T. H. V. and Rich, P. V. 2000. Dinosaurs of darkness. Indiana University Press, Bloomington. 222 p. Rich, T. H., Vickers-Rich, P., & Gangloff, R. A. 2002. Polar dinosaurs. Science 295 (5557): 979-980. Sachs, S. & Grant-Mackie, J. A. 2003. An ichthyosaur fragment from the Cretaceous of Northland, New Zealand. Journal of the Royal Society of NZ 33 (1): 307-314. Schellart, W. P., Lister, G. S., and Toy, V. G. 2006. A Late Cretaceous and Cenozoic reconstruction of the Southwest Pacific region: tectonics controlled by subduction and slab rollback processes. Earth-Science Reviews 76: 191-233. Stace, G. & Eagle, M. K. 2001. Yes! We had dinosaurs. Penguin, Auckland. 80 p. Stilwell, J. D. 1997. Tectonic and palaeobiogeographic significance of the Chatham Islands, South Pacific, Late Cretaceous fauna. Palaeogeography Palaeoclimatology Palaeoecology 136 (1-4): 97-119. Stilwell, J. D. & Zinsmeister, W. J. 1992. Molluscan systematics and biostratigraphy of the Lower Tertiary La Meseta Formation, Seymour Island, Antarctic Peninsula. American Geophysical Union, Antarctic Research Series 55: 1-192. Stilwell, J. D., Consoli, C. P., Sutherland, R., Salisbury, S., Rich, T. H., Vickers-Rich, P. A., Currie, P. J., & Wilson, G. J. 2006. Dinosaur sanctuary on the Chatham Islands, Southwest Pacific: first record of theropods from the K-T boundary Takatika Grit. Palaeogeography Palaeoclimatology Palaeoecology 230 (3-4): 243-250. Weishampel, D. B., Dodson, P., & Osmólska, H. 2004. (eds), The Dinosauria. University of California Press, Berkeley. 861 p. Welles, S. P. & Gregg, D. R. 1971. Late Cretaceous marine reptiles of New Zealand. Records of the Canterbury Museum 9: 1-111. Wiffen, J. 1981a. Moanasaurus, a new genus of marine reptile (Family Mosasauridae) from the Upper Cretaceous of North Island, New Zealand. NZ journal of geology and geophysics 23: 507-528. Wiffen, J. 1981b. The first Late Cretaceous turtles from New Zealand. NZ journal of geology and geophysics 24 (2): 292-300. Wiffen, J. 1990. New mosasaurs (Reptilia: Family Mosasauridae) from the Upper Cretaceous of North Island, New Zealand. NZ journal of geology and geophysics 33: 67-85. Wiffen, J. 1996. Dinosaurian palaeobiology: a New Zealand perspective. Memoirs of the Queensland Museum 39 (3): 725-731. Wiffen, J. & Moisley, W. L. 1986. Late Cretaceous reptiles (families Elasmosauridae, Pliosauridae) from the Mangahouanga Stream, North Island, New Zealand. New Zealand journal of geology and geophysics 29: 205-252. Wiffen, J. & Molnar, R. E. 1988. First pterosaur from New Zealand. Alcheringa 12: 53-59. Wilson, G. J., Schiøler, P., Hiller, N., & Jones, C. M. 2005. Age and provenance of Cretaceous marine reptiles from the South Island and Chatham Islands, New Zealand. NZ Journal of Geology and Geophysics 48 (2): 377-387. CAMPBELL H.J. (1996).- Titahi Bay fossil forest floor.- Geological Society of New Zealand, Newsletter, Auckland, n° 110, p. 22-24. CAMPBELL H.J. (1997).- Wellington fossil localities. Geological Society of New Zealand Annual Conference Field Trip Guide FT 1.- Geological Society of New Zealand, Miscellaneous Publication Series, Auckland, MP95B, p. 1-17. COOPER R.A. (ed., 2004).- The New Zealand geological timescale.- Institute of Geological and Nuclear Sciences, Monograph, Lower Hutt, n° 22, 284 p. CRUICKSHANK A.R.I. & FORDYCE R.E. (2002).- A new marine reptile (Sauropterygia) from New Zealand: further evidence for a Late Cretaceous Austral radiation of cryptoclidid plesiosaurs.- Palaeontology, London, vol. 45, n° 3, p. 557-575. EDWARDS A.R. (1991).- The Oamaru Diatomite.- New Zealand Geological Survey, Paleontological Bulletin, Wellington, n° 64, 260 p. FINLAY H.J. & MARWICK J. (1947).- New divisions of the New Zealand Upper Cretaceous and Tertiary.- New Zealand Journal of Science and Technology, Wellington, (B, General research section), vol. 28, p. 228-236. FLEMING C.A. (1979).- The Geological history of New Zealand and its life.- Auckland University Press, Auckland, 141 p. FORDYCE R.E. (2002).- Vanished world fossil trail, North Otago, New Zealand.- Vanished World Inc., Duntroon, pamphlet. FORDYCE R.E. & JONES C.M. (1990).- The history of penguins, and new fossil penguin material from New Zealand. In: DAVIS L.S. & DARBY J.D. (eds.), Penguin biology.- Academic Press, San Diego, p. 419-446 HAYWARD B.W. (1988).- Clifden Scientific Reserve, Southland.- Geological Society of New Zealand, Newsletter, Auckland, n° 80, p. 16-21. HAYWARD B.W. (1990).- Trilobites, dinosaurs and moa bones. The story of New Zealand fossils.- Bush Press, Auckland, 104 p. HAYWARD B.W. & WARD B. (1987, unpublished).- Inventory of New Zealand fossil localities of international, national and regional importance.- Geological Society of New Zealand, Auckland, Unpublished Report 87/1, 106 p. HORNIBROOK N. de B. (1990).- Hutchinsons Quarry and Target Gully shell pit geological reserves, Oamaru.- Geological Society of New Zealand, Newsletter, Auckland, n° 89, p. 46-49. KEYES I.W. (1984).- Joan WIFFEN and the Mangahouanga Stream reptiles: a profile.- Geological Society of New Zealand, Newsletter, Auckland, n° 63, p. 29-32. LEWIS D.W. (1984).- Chancet Rock Scientific Reserve.- Geological Society of New Zealand, Newsletter, Auckland, n° 66, p. 61-63. POLE M. (1990).- The Curio Bay Scientific Reserve.- Geological Society of New Zealand, Newsletter, Auckland, n° 89, p. 34-37. WEAVER S.D. & HAYWARD B.W. (1983).- An inventory and assessment of New Zealand geological and geomorphological features of national and regional importance.- Geological Society of New Zealand, Newsletter, Auckland, n° 59, p. 22-24. Australie Chinsamy, A., T.H.Rich & P.Vickers-Rich 1998 Polar dinosaur bone histology. Journal of Vertebrate Paleotology18:385 Constantine, A., A.Chinsamy, T.H.Rich, & P.Vickers-Rich. 1998. Periglacial environments and polar dinosaurs. South African Journal of Science 94:137-141. Douglas, J.G., & G.E.Williams. 1982. Southern polar forests: The Early Cretaceous floras of Victoria and their palaeoclimatic significance. Palaeogeography, Palaeoclimatology, Palaeoecology39:171-185. Martin, A.J. 2009 Dinosaur burrows in the Otway Group (Albian) of Victoria, Australia, and their relation to Cretaceous polar environments. Cretaceous Research 30(5):1223-1237 (PDF) Parrish, J.T., R.A.Spicer, J.G.Douglas, T.H.Rich, & P.Vickers-Rich. 1991. Continental climate near the Albian South Pole and comparison with climate near the North Pole. Geological Society of America, Abstracts with Programs 23:A302. Rich, T.H. 1996 Significance of polar dinosaurs in Gondwana. Memoirs of the Queensland Museum 39:711-717. Rich, P.V. and T.H.Rich 1999 Wildlife of Gondwana. The 500-million year history of vertebrate animals from the ancient southern supercontinent. Second Edition. Reed books, Sydney. Rich, P.V., L.S.Rich and T.H.Rich 1996 Australia's Lost World. A history of Australia's backboned animals. Kangaroo Press, Monash Science Centre, Monash University. Rich, P.V., Rich, T.H., Wagstaff, B.E., McEwen-Mason, J., Douthitt, C.B., Gregory, R.T. and Felton, E.A. 1988. Evidence for low temperatures and biologic diversity in Cretaceous high latitudes of Australia. Science 242:1403-1406. Rich, T.H., Rich, P.V., Wagstaff, B., McEwen-Mason, J., Douthitt, C.B. and Gregory, R.T. 1989. Early Cretaceous biota from the northern side of the Australo-Antarctic Rift Valley. In J.A.Crame (ed.) Origins and Evolution of the Antarctic Biota. Geological Society Special Publication 47:121-130. Rich, T.H., R.A.Gangloff, & W.Hammer. 1997. Polar Dinosaurs. In P.J. Currie & K.Padian (eds.), Encyclopedia of Dinosaurs, pp. 562-573. Academic Press. Rich, T.H. and P.V.Rich 1988. A juvenile dinosaur brain from Australia. National Geographic Research. 4(2):148. Rich, T.H. and P.V.Rich 1989 Polar dinosaurs and biotas of the Early Cretaceous of southeastern Australia. National Geographic Research 5:15-53. Rich,T.H., P.Vickers-Rich & R.A.Gangloff 2002 Polar Dinosaurs. Science 295:979-980 Vickers-Rich, P., T.H.Rich 2000 Dinosaurs of Darkness. Indiana University Press<============= Un Theropode en mode Harfang Un Ceratopsien des neige
-
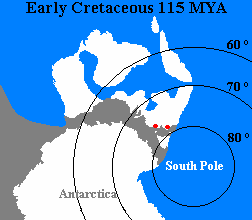
pour le volcanisme au Crétacé en New Zealand. Le gisement de dinosaure de New Zealand.C'est pas la profusion de dinos, par contre en reptile marin a long cou=>kaiwhekea katiki Au Crétacé, la New Zealand était dans le cercle polaire, il y avait une nuit polaire, une baisse des températures, Des études isotopiques de l'oxygène donnent une température annuelle moyenne de -2 ° +/- 5 ° Celsius et un arrêt de la production végétale et observable sur les ligne de croissance des tronc fossile . La flore, Les fossiles donne l'image d'un environnement forestier polaire mais bien végétalisé sans équivalent actuel, dominé par des fougères, des gingko et des conifères, Il est envisager que certains dinosaures du godwana migrer, que d'autre hiberné comme le timimus ou creusaient des terriers( dinosaur polar.). Il y a beaucoup plus de chose en Australie.
-
On connait la musique, C'est comme çà!
-
Salut Chef! Il faut que je développeQuoi?
-
En quartzite! et creusse dans le permafrost(coucou!)C'est tradition acheuléen!
-
L'ambre & les résines fossiles
gaeldeploeg a répondu à un sujet dans Forum Fossiles et Paléontologie
Le terme usuel ou vernaculaire pour parlé d'une résine fossile, c'est ambre. Point barre. -
Annonce importante pour ceux qui recherche le Lacroix
gaeldeploeg a répondu à un sujet dans Echanges de minéraux
Oui le sablais, mais=> il y a les autres aussi Un clique sur les liens en bleu pour ouvrir Mineralogie de la France et de ses colonies : description physique et chimique des mineraux, etude des conditions geologiques de leurs gisements, tome quatrieme by Lacroix, Alfred, 1863-1948 Published 1910 Topics Mineralogy--France, Mineralogy--France--Colonies SHOW MORE Volume 1, 2, 3, 4 & 5. "This monumental work describes the minerals of France and its colonies, including Madagascar; numerous gemstones are included, especially from Madagascar, but more complete information is found in Lacroix's work on them minerals of Madagascar ... Among many topographical mineralogies that have been published this work stands without peer, being extremely thorough, neglecting no important aspect, and lavishly illustrated."--Gemology by J. Sinkankas, entry no. 3736 Publisher Librarie Polytechnique, Paris Year 1910 Language French Collection gialibrary; additional_collections Notes Page 643 is misnumbered 634. Mineralogie de la France et de ses colonies : description physique et chimique des mineraux, etude des conditions geologiques de leurs gisements, tome premier by Lacroix, Alfred, 1863-1948 texts eye100 favorite0 comment0 GIA Mineralogie de la France et de ses colonies : description physique et chimique des mineraux, etude des conditions geologiques de leurs gisements, tome cinquieme by Lacroix, Alfred, 1863-1948 texts eye344 favorite0 comment0 GIA Mineralogie de la France et de ses colonies : description physique et chimique des mineraux, etude des conditions geologiques de leurs gisements, tome deuxieme by Lacroix, Alfred, 1863-1948 texts eye88 favorite0 comment0 GIA Mineralogie de la France et de ses colonies : description physique et chimique des mineraux, etude des conditions geologiques de leurs gisements, tome troisieme by Lacroix, Alfred, 1863-1948 texts eye89 favorite0 comment0 GIA Mineralogie de Madagascar, tome 2 by Lacroix, Alfred, 1863-1948 texts eye169 favorite0 comment0 GIA Mineralogie de Madagascar, tome 1 by Lacroix, Alfred, 1863-1948 texts eye209 favorite0 comment0 -
Annonce importante pour ceux qui recherche le Lacroix
gaeldeploeg a répondu à un sujet dans Echanges de minéraux
Un gratos en version virtuelhttps://archive.org/stream/MineralogieDeLaFranceEtDeSesColoniesDescriptionPhysiqueEtChimique_60/lacroix-a-france-vol4-1910-RTL014357-LowRes#page/n667/mode/2up -
Par ce que j'en ai marre d'entendre et de lire des connerie. Le "Mineralogie de la France et de ses colonies". https://archive.org/stream/MineralogieDeLaFranceEtDeSesColoniesDescriptionPhysiqueEtChimique_60/lacroix-a-france-vol4-1910-RTL014357-LowRes#page/n667/mode/2up
-
L'ambre & les résines fossiles
gaeldeploeg a répondu à un sujet dans Forum Fossiles et Paléontologie
https://archive.org/stream/MineralogieDeLaFranceEtDeSesColoniesDescriptionPhysiqueEtChimique_60/lacroix-a-france-vol4-1910-RTL014357-LowRes#page/n667/mode/2up Le Lacroix, Mineralogie de la France et de ses colonies. Donc, La nomenclature des résines fossiles et simple!
-
Oui, là!http://planet-terre.ens-lyon.fr/image-de-la-semaine/Img322-2010-08-30.xmlentre autre.
-
C'est Nel. A. échantillons de Fourtou, Aude. Cénomanien marin. Arbre producteur: Araucaria. Ah, Merci à ......., membre de ce forum Girard_etal_2013_Fourtou-amber.pdf et Pour avoir une idée sur l'ampleur du gisement a résine fossile Cénomaniene. ( La honte) La cause d'une telle production: et le stresse sodique inhérent et la fréquence des inclusions d'origine marine dans cette ambre!